4.2. DIATOMEAS Y MEDIO AMBIENTE MARINO (I)
Otras investigaciones más detalladas proporcionan datos particularmente interesantes al efectuarse selectivamente, primero sobre organismos animales y después sobre organismos vegetales, finalizando por los de transición.
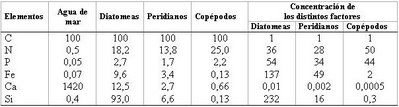
Cuadro 5.1. Composición relativa de los organismos planctónicos (con relación a C = 100)
Tomado de AUBERT, Maurice. El cultivo del océano. Labor. Barcelona, 1968.
Desde 1890 se sabe que el plancton, y por tanto las diatomeas, pueden cultivarse. Se conocen los medios de hacer que se reproduzca el fitoplancton, y, a partir de éste, de alimentar el zooplancton en cría artificial.
Lo anterior significa que el crecimiento de las diatomeas está determinado por las condiciones medioambientales.
Señalaremos ahora las condiciones y las dificultades de este cultivo y de esta cría de tipo tan particular.
Hay que tener presente, desde un buen principio, que las condiciones vitales creadas de nuevo en una probeta son forzosamente en extremo distintas de las que existen en el medio marino. Por lo tanto, la dificultad consiste en determinar las condiciones del medio natural y adaptarse a ellas con la mayor fidelidad posible.
Los principales ensayos de cultivo se han efectuado con algas planctónicas, diatomeas y peredinios. Su crecimiento y producción resultan favorecidos por factores que pueden variar conjuntamente o independientemente unos de otros. En las condiciones naturales, cada especie tiene una zona de temperatura preferente, pero los numerosos estudios in vitro efectuados en este terreno, como el de Harvey, que data de 1939, y el de Jorgensen, de 1960, han demostrado que la influencia de la temperatura en el crecimiento de una especie en cultivo depende asimismo de los demás factores ambientales, y, en particular, de la luz.
Señalemos el ejemplo de la Nitzschia ovalis: a 1500 lx (lux) el crecimiento máximo se efectúa entre 16 y 18 ºC; para 2250 lx, este punto óptimo del crecimiento se encuentra en niveles térmicos diferentes, entre 14 y 18 ºC por una parte, y por otra, entre 27 y 29 ºC. A 6500 lx, el crecimiento más rápido se verifica entre 27 y 30 ºC; más allá, las temperaturas ya no son soportables para esta especie.
Tenemos, pues, un ejemplo de la acción ejercida por dos factores específicos del medio ambiente, pero veremos que la complejidad va en aumento a medida que profundizamos en este nuevo problema. Los trabajos de Hutner y sus colaboradores han demostrado que una de las acciones de la temperatura consistía en aumentar las necesidades nutritivas. Por ejemplo, para Euglena, aumenta la necesidad de vitaminas B12 con el aumento de la temperatura, y este efecto nutritivo de orden térmico puede tener importancia ecológica: las formas de aguas frías se vuelven entonces capaces de colonizar aguas cálidas, si éstas se hallan enriquecidas.
El umbral de intensidad luminosa, sin embargo, difiere según las especies en cultivo, ya porque ésta se proporcione de manera continua, o bien durante períodos más o menos largos de iluminación. Algunos cultivos de diatomeas, como Chaetoceros didimus, a 19 ºC requieren una iluminación de doce horas diarias, con una intensidad de 200 bujías por pie cuadrado.
Es de sobra conocida la importancia biológica que tiene el complejo temperatura-salinidad sobre el medio marino. No es raro que este segundo factor interfiera directamente en la evolución de nuestros cultivos planctónicos. Ryther, Droop y Guillard han demostrado la importancia que esto tiene. En el medio natural, la salinidad varia geográficamente según las profundidades, el alejamiento de las costas, etc., y así las diferentes especies se hallan divididas selectivamente por fronteras completamente teóricas, es cierto, pero sin embargo reales. Otras, menos sensibles a estas diferencias salinas, pueden franquear estos limites. Ryther y Guiliard han estudiado la influencia de la salinidad en particular sobre Cyclotella nana, demostrando que el índice de crecimiento de las cepas de estuario es invariable, aunque la salinidad varíe desde el valor acostumbrado del agua normal (3,5 %) hasta los valores del agua casi dulce (0,05 %).
Asimismo, el índice de crecimiento de las cepas aisladas del agua de la plataforma continental disminuye rápidamente si la salinidad se hace inferior a la mitad del valor del agua de mar normal. Parece ser, pues, que la tolerancia a ciertos iones, y en particular al ion sodio, es más o menos extensa según las especies.
Pero no basta con poner a los seres que deseamos cultivar en un medio que no les resulte mortal o que les sea más o menos favorable; hay que pensar también en alimentarlos, y entonces entra en juego la importancia de los factores nutritivos. Como sabemos, el agua de mar contiene todos los elementos conocidos en mayor o menor disolución, y sabemos también que los organismos tienen necesidad de los elementos principales, como el carbono, el hidrógeno, el oxigeno, el nitrógeno, el fósforo, el potasio, el azufre, el sodio, el magnesio y el calcio. También tienen necesidad de elementos menores, que utilizan en estado de vestigios, como son el hierro, el cinc, el manganeso, el cobalto, el vanadio, el cobre, etc.
Por lo tanto, es preciso suministrar todos estos elementos de manera asimilable a dichos cultivos; les podemos dar agua de mar enriquecida con extractos terrígenos; un medio nutritivo adecuado como el de Erdschreiber, aporta vestigios de metales y factores de crecimiento. Se puede utilizar asimismo un medio totalmente artificial, lo cual es posible después de los estudios de Provasoli, Laughlin, Droop, Chu y Rodhe que, estudiando sistemáticamente cada elemento nutritivo, lograron preparar medios específicos dotados de un valor de crecimiento óptimo para cada variedad de organismo. Así fue como Provasoli, Laughijn y Droop lograron preparar un agua de mar artificial, químicamente definida que se puede esterilizar sin provocar precipitado a causa de la disminución del porcentaje de calcio y del aumento del de magnesio. Los fosfatos han sido reemplazados por glicerofosfatos, y, por otra parte, estos autores añaden agentes quelantes que sirven para estabilizar los oligoelementos, formando complejos fácilmente reversibles con estos metales. El más empleado es el E. D. T. A. (ácido etilendiamino-tetraacético), que es metabólicamente casi inerte para los organismos y que, a causa de ello, no ayuda al crecimiento de las contaminaciones en los cultivos no estériles. Estos oligoelementos, por otra parte, sólo se utilizan a concentraciones del orden del microgramo por litro.
Pero además de estos factores que favorecen el crecimiento y que sólo se utilizan en forma de vestigios, existen factores nutritivos esenciales, a los que ahora vamos a pasar revista. Una necesidad esencial es la del nitrógeno. Según Myers, la mayoría de las algas unicelulares contienen aproximadamente 8 % de nitrógeno; luego cada gramo por litro (en peso seco) de células producidas en un cultivo, requiere 80 mg/l de nitrógeno, o sea 580 mg/l de nitrato potásico, forma en la cual se suministra. Las fuentes de nitrógeno, por otra parte, pueden variar, pudiéndose utilizar el amoniaco, el nitrato, el nitrito y la urea, pues cada especie utiliza con mayor o menor rapidez una u otra de estas fuentes. Es preferible utilizar el nitrato cuando aumenta el pH del medio ambiente, y, en cambio, el amoniaco si el pH disminuye.
El segundo factor nutritivo importante: el fósforo. En la naturaleza, su índice constituye en general uno de los factores limitadores del crecimiento (ésta es, por otra parte, la razón de que cuando se quieren enriquecer fiordos o lagunas, se enriquecen con superfosfatos). El papel energético y sintético de este elemento es bien conocido en bioquímica, aunque su acción en el medio acuático no esté aún totalmente elucidada por lo que a su mecanismo se refiere. Las experiencias de Kuenzier y Ketchum en 1962 demostraron que las diatomeas no se dividían si el contenido en fósforo de las células era inferior al valor mínimo de 6x10-14 g por célula, dirigiéndose a Phaeodactylum pero estas células pueden acumular hasta treinta veces esta cantidad en presencia de un exceso y dividirse después a expensas de la reserva citada, cuando haya falta de fósforo en el ambiente.
La tercera fuente de nutrición esencial: el carbono. Todas las células necesitan carbono, que se incorpora mediante el mecanismo de la fotosíntesis con utilización del gas carb6nico, o bien explotando las fermentaciones y utilizando entonces moléculas más complejas.
En los cultivos in vitro hay que utilizar fuentes variadas según las especies: glucosa, lactato, ácido glutámico o málico, glicerol, etc.
Se puede emplear igualmente el gas carbónico, fuente sencilla que es casi siempre la preferida. Myers indica que para obtener 1 mg de alga en peso seco, se requieren 0,5 mg de carbono, o sea 1,8 mg de CO2 En la práctica basta con inyectar en el medio elegido aire enriquecido con CO2 a 5 %, que permita realizar al mismo tiempo la ventilación y la agitación del medio, pero como los organismos cultivados sacan toda su energía de reacción de oxidorreducción, se requiere un receptor de hidrógeno (o un donante de electrones). Por lo tanto, la ventilación proporciona este receptor en forma de oxígeno, cuyo porcentaje en el aire ambiente es suficientemente elevado.
Finalmente, para que el mecanismo nutritivo pueda efectuarse de manera conveniente, es necesaria una aportación vitamínica suficiente, pues la mayoría de seres vivientes no efectúan por sí mismos. La síntesis de las vitaminas que les son necesarias empíricamente.


Comentarios